|
План:
1. Статистичний характер законів успадкування.
2. Відхилення при розщепленні.
3. Зчеплене успадкування.
Література: Загальна біологія: Підруч для 11 кл./ М.Є. Кучеренко та інш. 2006. с. 49-58
1. На той час, коли Г. Мендель проводив свої досліди з горохом, ще нічого не було відомо про гени, будову хромосом і процес мейозу. Але він запропонував закон чистоти гамет, який згодом знайшов своє експериментальне підтвердження. Цей закон стверджує, що в гібридного (гетерозиготного) організму гамети «чисті». Тобто кожна з гамет такого диплоїдного організму може мати лише один алельний ген і не може одночасно нести дві алелі.
Нестатеві (соматичні) клітини, як правило, мають диплоїдний набір хромосом, тобто в кожній з них є два алельні гени. Це можуть бути дві домінантні чи дві рецесивні алелі певного гена (гомозигота) або домінантна і рецесивна алелі (гетерозигота). Коли внаслідок мейотичного поділу утворюються статеві клітини, в кожну з них потрапляє тільки одна алель з кожної пари. Гомозиготна особина формує лише один сорт статевих клітин (домінантною або рецесивною алеллю певного гена), тоді як гетерозиготна - два сорти в рівних кількостях (50% з домінантною алеллю певного гена і 50% — з рецесивною).
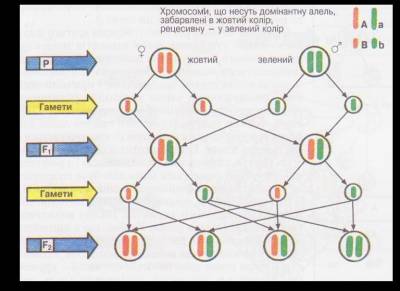
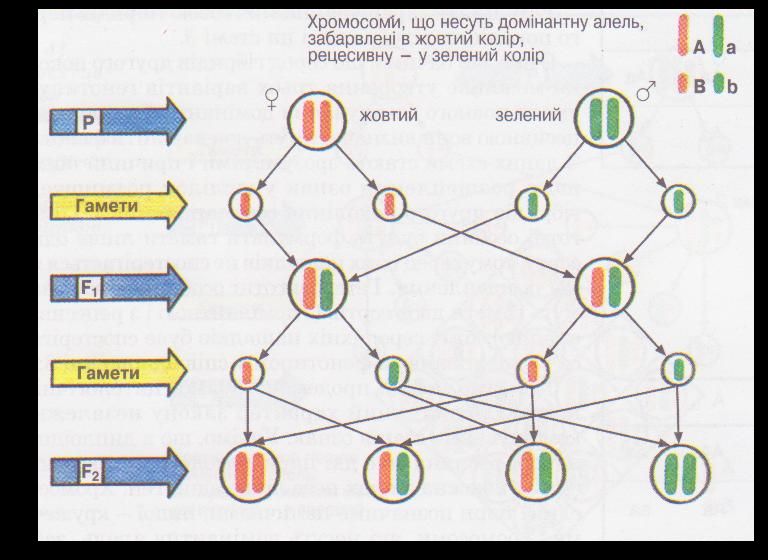
Мал. 1. Цитологічні основи розщеплення при моно гібридному схрещуванні.
Користуючись малюнком 1, можна простежити за гомологічними хромосомами при моногібридному схрещуванні гомозиготних особин гороху посівного. Для спрощення припустимо, що такі особини мають лише одну пару гомологічних хромосом (тобто кількість хромосом у диплоїдному наборі дорівнює двом: 2п = 2), а кожна з них містить тільки один ген. Хромосому з домінантною алеллю (А) на малюнку позначено жовтим кольором, а з рецесивною (а) — зеленим. Відомо, що нащадки, отримані від схрещування особин гомозиготних за домінантною та рецесивною алелями (гібриди першого покоління), будуть гетерозиготними (їхній генотип — Аа). Це пояснюється тим, що одну хромосому з домінантною алеллю вони дістають від одного з батьків, а іншу, з рецесивною, — від другого. Отже, такі рослини будуть одноманітними як за генотипом, так і за фенотипом.
Гібриди першого покоління, на відміну від батьків, утворюватимуть гамети двох сортів: половина з них нестиме хромосому з домінантною алеллю, половина - з рецесивною. Внаслідок схрещування гібридів першого покоління між собою у їхніх нащадків (гібридів другого покоління) можливі три варіанти генотипів: чверть особин матиме хромосоми лише з домінантними (гомозиготи за домінантною алеллю - АА), половина - одну хромосому з домінантною, другу - з рецесивною (гетерозиготи - Аа) і чверть - хромосоми лише з рецесивними (гомозиготи за рецесивною алеллю - аа) алелями. А за фенотипом три чверті насіння гібридів другого покоління матиме жовте забарвлення (гомозиготи за домінантною алеллю і гетерозиготи), а одна чверть — зелене (гомозиготи за рецесивною алеллю).
Отже, якщо утворюється значна кількість гамет рівної життєздатності, то стає зрозумілим статистичний характер закону розщеплення. Він визначається рівною ймовірністю зустрічей гамет різних сортів. Результати проведеного Г. Менделем моногібридного схрещування можна записати схематично (схема 1).
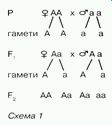
Проте простіше записати хід схрещування за допомогою так званої решітки Пеннета. її зображують так: по горизонталі записують гамети однієї (у роздільностатевих організмів - батьківської) особини, а по вертикалі — іншої (материнської). В місцях перетину горизонтальної та вертикальної ліній зазначають генотипи нащадків (схема 2).
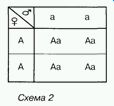
Згідно з записом, усі нащадки, одержані від схрещування особин, гомозиготних за домінантною і рецесивною алелями, є одноманітними й гетерозиготними.
Результати схрещування між собою гібридів першого покоління відображені на схемі 3.
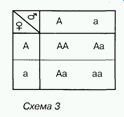
Отже, серед гібридів другого покоління можливе утворення трьох варіантів генотипу. За умови повного домінування домінантної алелі над рецесивною вони визначатимуть два варіанти фенотипу. З даних схеми стають зрозумілими і причини подальшого розщеплення ознак унаслідок розмноження гібридів другого покоління самозапиленням. Гомозиготні особини будуть формувати гамети лише одного сорту, тому серед їхніх нащадків не спостерігається явище розщеплення. Гетерозиготні особини утворюватимуть гамети двох сортів (з домінантною і з рецесивною алеллю), тому серед їхніх нащадків буде спостерігатися розщеплення за фенотипом у співвідношенні 3:1.
Так само можна продемонструвати цитологічні основи і статистичний характер закону незалежного комбінування станів ознак. Уявімо, що в диплоїдному наборі рослина має дві пари гомологічних хромосом (2п = 4), кожна з яких несе лише один ген. Хромосоми однієї пари позначимо паличками, іншої — кружечками. Хромосоми, які несуть домінантну алель, забарвлені у жовтий колір, а рецесивну — в зелений.
Припустімо, що материнський організм містить хромосоми лише з домінантними алелями генів забарвлення і структури поверхні насіння, а батьківський - лише з відповідними рецесивними. Такі гомозиготні організми в процесі утворення гамет формуватимуть один їхній сорт - або з домінантними, або з рецесивними алелями. У разі схрещування батьківських форм усі нащадки (гібриди першого покоління) дістають по дві хромосоми з відповідними домінантними алелями від материнського організму і по дві з рецесивними — від батьківського. Тож усі вони будуть гетерозиготними за генами забарвлення і структури поверхні насіння й формуватимуть чотири сорти гамет у рівних кількостях. При цьому в двох з них алельні гени будуть знаходитися у тих самих комбінаціях, що і в гаметах вихідних батьківських форм, а в двох інших — у нових поєднаннях (рекомбінаціях). Рекомбінація (від лат. ре - префікс, який означає поновлення, повтор дій, і комбінатів — поєднання) — перерозподіл спадкового матеріалу батьків у генотипі нащадків. Іншими словами, рекомбінації — це нові поєднання алелей різних генів у гаметах гібридів, які відрізняються від їхнього поєднання у гаметах батьків.
Унаслідок рівної ймовірності зустрічей гамет різних сортів, які утворюють гібриди першого покоління, в їхніх нащадків (гібридів другого покоління) можливе утворення дев'яти варіантів генотипів. Ці варіанти, у свою чергу, визначатимуть чотири різних варіанти фенотипу. У двох із них стани ознак забарвлення насінини і структури її поверхні будуть у тих самих поєднаннях, як і у фенотипі вихідних батьківських форм (жовтий колір — гладенька поверхня насінини і зелений колір — зморшкувата поверхня), а у двох — в нових — рекомбінаціях (жовтий колір — зморшкувата поверхня і зелений колір — гладенька поверхня). Рекомбінації е одним із джерел мінливості організмів.
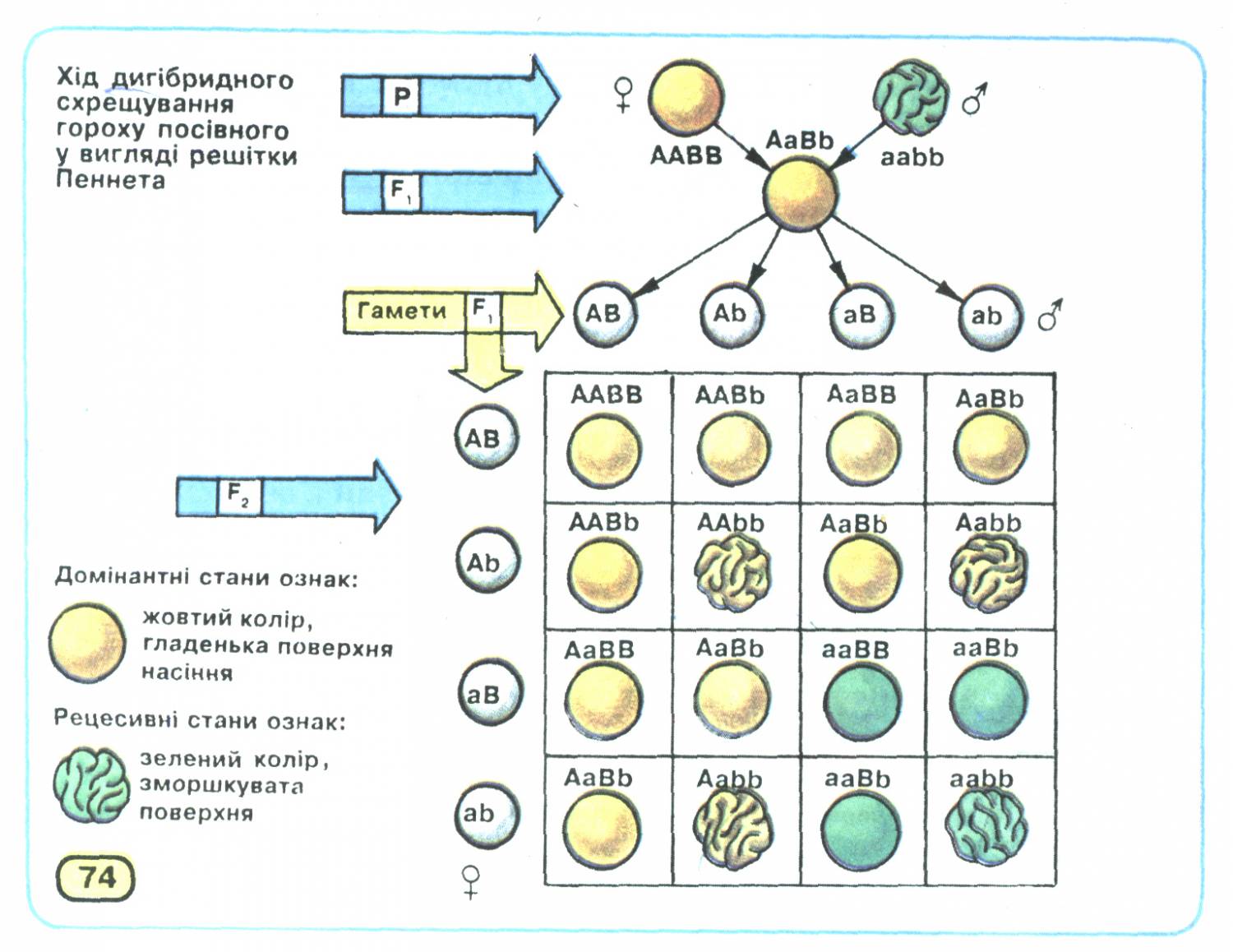
Статистичний характер закону незалежного комбінування станів ознак можна довести, записавши хід дигібридного схрещування за допомогою решітки Пеннета. При схрещуванні гомозиготних батьківських форм його результат буде такий:
Результати схрещування батьківських форм, гомозиготних за домінантними і відповідно рецесивними алелями генів забарвлення (А - а) і структури поверхні (В — Ь) насіння, розташованих у негомологічних хромосомах, підтверджують закон одноманітності гібридів першого покоління: всі вони гетерозиготні за цими генами й утворюватимуть насіння жовтого кольору з гладенькою поверхнею.
Хід схрещування гібридів першого покоління між собою наведено на малюнку 2. Він свідчить про те, що серед гібридів другого покоління розщеплення за фенотипом буде таким: 9 частин насіння жовтого кольору з гладенькою поверхнею, 3 — жовтого зі зморшкуватою, 3 — зеленого з гладенькою і 1 — зеленого зі зморшкуватою. Отже, розщеплення серед гібридів другого покоління як за ознакою забарвлення насіння, так і за ознакою структури його поверхні становитиме 12:4 (тобто 3:1). За умови повного домінування домінантних алелей над відповідними рецесивними фенотип жовтого насіння з гладенькою поверхнею визначатиметься чотирма варіантами генотипу (ААВВ, ААВЬ, АаВВ, АаВЬ), жовтого зі зморшкуватою - двома (AAbb, Aabb), зеленого з гладенькою - теж двома (ааВВ, ааВЬ), а зеленого зі зморшкуватою - одним (aabb).
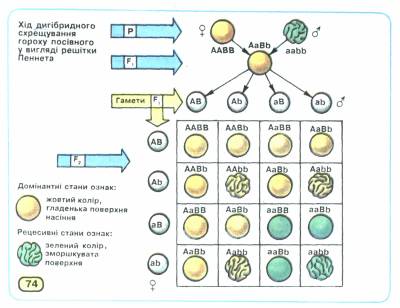
Мал. 2. Хід дигібридного схрещування гороху посівного.
2. Як відомо, багато законів має винятки.
Розглядаючи цитологічні основи законів спадковості та їхній статистичний характер, вважалося, що один із алельних генів повністю пригнічує прояв іншого у фенотипі гібридної особини. Внаслідок цього у гетерозиготних організмів проявляється лише один із двох протилежних станів певної ознаки (домінантний). Але є домінантні алелі, які лише частково переважають над рецесивними. У таких випадках гетерозиготна особина за фенотипом більше нагадуватиме особину, гомозиготну за домінантною алеллю, але дещо відрізнятиметься від неї. Таке явище називають неповним домінуванням.
Трапляються випадки, коли жодна з алелей не домінує над іншою. Тоді спостерігають проміжний характер успадкування. В результаті схрещування гомозиготних рослин нічної красуні, одна з яких мала червоне забарвлення віночка квітки, а інша – біле, отримані гібриди першого покоління хоча і були одноманітними, але їхній віночок був забарвлений у рожевий колір, тобто займав наче проміжне положення між відповідними фенотипами батьківських особин.
Схрещуючи гібриди першого покоління між собою, спостерігали таке розщеплення серед їхніх нащадків за фенотипом: одна чверть гібридів другого покоління мала червоне забарвлення віночка, половина — рожеве і ще одна чверть — біле. Отже, через те, що жодна з алелей не домінує над іншою, фенотип гетерозиготних особин займає наче проміжне положення між відповідними фенотипами гомозиготних батьків, а серед гібридів другого покоління розщеплення за фенотипом відбувається у співвідношенні 1:2:1, а не 3:1, якби одна алель повністю домінувала над іншою.
Отже, у разі проміжного характеру успадкування у гібридів другого покоління можливі три варіанти генотипу, кожен з яких визначатиме свій варіант фенотипу.
У разі повного домінування однієї алелі над іншою особини, гомозиготні за домінантною алеллю і гетерозиготні, подібні за фенотипом, їхній генотип можна визначити за фенотипом нащадків, одержаних від певних типів схрещувань, наприклад за допомогою аналізуючого схрещування. Аналізуюче схрещування — це схрещування особини, генотип якої хочуть визначити, з особиною, гомозиготною за рецесивною алеллю досліджуваного гена. Таке схрещування ґрунтується на тому, що особини, гомозиготні за рецесивною алеллю певного гена, завжди мають лише певний варіант фенотипу й утворюють гамети лише одного сорту.
Отже, якщо серед нащадків, одержаних від аналізуючого схрещування, розщеплення за фенотипом не спостерігається, то батьківська особина, генотип якої перевіряли, була гомозиготною за домінантною алеллю.
Якщо внаслідок аналізуючого схрещування серед нащадків спостерігатиметься розщеплення за фенотипом у співвідношенні 1:1, то особина, чий генотип визначали — гетерозиготна за досліджуваним геном.
Аналізуюче схрещування широко застосовують у селекційній роботі, оскільки воно дає можливість визначити генотип особин, яких планують використати для одержання нащадків із певним фенотипом. Зокрема, завдяки цьому можна уникнути небажаного розщеплення за фенотипом серед нащадків, усунувши від розмноження гетерозиготних особин.
Відхилення від очікуваних результатів розщеплення часто пов'язані з дією так званих летальних алелей, оскільки особини, гомозиготні й гетерозиготні за цими алелями, мають різну життєздатність.
Летальні алелі (від лат. леталіс — смертельний) — це алелі, які, проявляючись у фенотипі, спричиняють загибель організму на тій чи іншій стадії його розвитку.
Оскільки летальні алелі можуть призводити до загибелі нащадків ще до народження, кількісне співвідношення різних фенотипних груп нащадків відрізнятиметься від того, яке можна було б очікувати теоретично, виходячи із закономірностей спадковості, встановлених Г. Менделем. Найчастіше летальні алелі рецесивні, тому вони можуть проявлятися у фенотипі лише у гомозиготному стані (наприклад, алель, яка спричиняє водянку в телят йоркширської породи).
3. Закон незалежного комбінування станів ознак ґрунтується на таких положеннях:
— розвиток різних станів ознак зумовлений алель-ними генами, які мають однакове розташування в гомологічних хромосомах;
— гамети та інші гаплоїдні клітини, які мають по одній хромосомі з кожної пари гомологічних хромосом, несуть лише один алельний ген із певної їхньої кількості;
— гени, що контролюють розвиток ознак, які успадковуються незалежно, розташовані в негомологічних хромосомах.
При доведенні статистичного характеру законів спадковості, встановлених Г. Менделем, то для спрощення припускалося, що кожна хромосома несе лише один ген. Але вчені вже давно звернули увагу на те, що кількість спадкових ознак організмів значно перевищує кількість їхніх хромосом у гаплоїдному наборі. Так, у гаплоїдному наборі мухи-дрозофіли, яка стала класичним об'єктом для генетичних досліджень, усього чотири хромосоми. Але кількість спадкових ознак і, відповідно, генів, які їх визначають, безсумнівно, значно більше.
Отже, в кожній хромосомі є не один, а багато генів. Разом з ознаками, які успадковуються незалежно, існують і такі, що успадковуються зчеплено одна з одною, оскільки визначаються генами, розташованими в одній хромосомі. Такі гени утворюють групу зчеплення. Кількість груп зчеплення в організмів певного виду дорівнює кількості хромосом у гаплоїдному наборі (наприклад, у дрозофіли 1n = 4, у людини -1n = 23).
Експериментальне явище зчепленого успадкування довів видатний американський генетик Т.Х. Морган зі своїми співробітниками.
Явище зчепленого успадкування Т.Х. Морган встановив за допомогою наступного досліду. Самців дрозофіли, гомозиготних за домінантними алелями забарвлення тіла (сіре) і форми крил (нормальна), він схрестив із самками, гомозиготними за відповідними рецесивними алелями (чорне забарвлення тіла — недорозвинені крила). Всі гібриди першого покоління мали сіре забарвлення тіла і крила нормальної форми, тобто були гетерозиготними за обома генами. Щоб з'ясувати генотип гібридів першого покоління, Т.Х. Морган провів аналізуюче схрещування: схрестив їх з особинами, гомозиготними за відповідними рецесивними алелями.
Теоретично від такого схрещування можна було очікувати два варіанти розщеплення. Якби гени, що визначають забарвлення тіла і форму крил, були розташовані в негомологічних хромосомах, тобто успадковувалися незалежно, розщеплення мало бути таким: 25% особин із сірим тілом і нормальною формою крил, 25% — із сірим тілом і недорозвиненими крилами, 25% — з чорним тілом і нормальними крилами і 25% — з чорним тілом і недорозвиненими крилами (тобто у співвідношенні 1:1:1:1). Але якби ці гени були розташовані в одній хромосомі та успадковувалися зчеплено, то серед нащадків можна було б очікувати 50% особин із сірим тілом і нормальною формою крил і 50% - з чорним тілом і недорозвиненими крилами (тобто у співвідношенні 1:1).
Реально 41,5% нащадків мали сіре тіло і нормальну форму крил, 41,5% - чорне тіло і недорозвинені крила, 8,5% - сіре тіло і недорозвинені крила і 8,5% - чорне тіло і нормальну форму крил. Отже, розщеплення за фенотипами наближувалося до 1:1 (як у випадку зчепленого успадкування). Але серед нащадків були й два інші варіанти фенотипів, які можна було б очікувати в тому випадку, якби гени забарвлення тіла і форми крил містились у негомологічних хромосомах й успадковувалися незалежно.
Т.Х. Морган припустив, що гени, які визначають забарвлення тіла мух і форму їхніх крил, розташовані в одній хромосомі й успадковуються зчеплено. Під час утворення статевих клітин у процесі мейозу гомологічні хромосоми здатні обмінюватися своїми ділянками (явище перехресту хромосом, або кросинговер). Як вам відомо, кросинговер підсилює мінливість, забезпечуючи утворення нових варіантів поєднань алелей. При цьому може відбуватись обмін кількома генами або ділянками одного гена.
Дослідження кросинговеру, проведені на різних організмах, виявили такі закономірності:
— сила зчеплення між двома генами, розташованими в одній хромосомі, обернено пропорційна відстані між ними. Отже, чим ця відстань більша, тим частіше відбувається кросинговер;
— частота кросинговеру між двома генами, розташованими в одній хромосомі, є величиною відносно постійною для кожної конкретної пари генів.
На підставі проведених досліджень зроблено важливий висновок - гени розташовані в хромосомі по її довжині в лінійному порядку, тобто один за одним. Згодом відкриття будови молекули ДНК підтвердило це. Як ви пам'ятаєте, в ній у лінійному порядку розташовані окремі мономери — нуклеотиди, якими закодовано спадкову інформацію.
Відносну сталість частоти кросинговеру між різними парами генів однієї групи зчеплення використовують як показник відстані між окремими генами, а також для визначення послідовності їхнього розташування в хромосомі. Зокрема, визначення частот кросинговеру між різними парами генів використовують для складання генетичних карт хромосом. На цих картах зазначається порядок розташування і відносні відстані між генами в певній хромосомі.
|